Beztlenowce to bakterie. Cechy beztlenowców, klasyfikacja i metody uprawy
Najbogatszą grupą organizmów pod względem liczby i różnorodności szlaków metabolicznych są prokarioty. Niektóre z nich używają schematu oddychania tlenowego typowego dla większości eukariontów do syntezy ATP (głównej energii "waluty" komórki). Mikroorganizmy, które nie posiadają tego mechanizmu, nazywane są beztlenowcami. Te bakterie są zdolne do wytwarzania energii ze związków chemicznych bez tlenu.
Klasyfikacja beztlenowa
W odniesieniu do tlenu rozróżnia się dwie grupy bakterii beztlenowych:
- opcjonalnie - może odbierać energię z lub bez tlenu, przejście z jednego rodzaju metabolizmu do innego zależy od warunków środowiskowych;
- obowiązkowe - nigdy nie używaj O 2 .
W przypadku beztlenowych beztlenowych metabolizm beztlenowy ma znaczenie adaptacyjne, a bakterie wykorzystują je tylko w ostateczności, gdy są uwalniane do środowiska beztlenowego. Wyjaśnia to fakt, że oddychanie tlenem jest o wiele bardziej korzystne energetycznie.
W drugiej grupie beztlenowców brakuje biochemicznego mechanizmu wykorzystywania O2 do utleniania związków, a obecność tego pierwiastka w środowisku jest nie tylko nie pomocna, ale także toksyczna.
Istnieje kilka rodzajów obligatoryjnych beztlenowców, różniących się odpornością na obecność tlenu cząsteczkowego:
- ciężka śmierć nawet przy niskich stężeniach O2;
- umiarkowanie ciężkie charakteryzujące się średnią lub wysoką odpornością na obecność tlenu;
- Aerotolerancyjny - specjalna grupa prokariotów, która może nie tylko przetrwać, ale także rozwijać się w powietrzu.
Stosunek konkretnej bakterii do tlenu można określić przez naturę jego wzrostu w grubości pożywki.
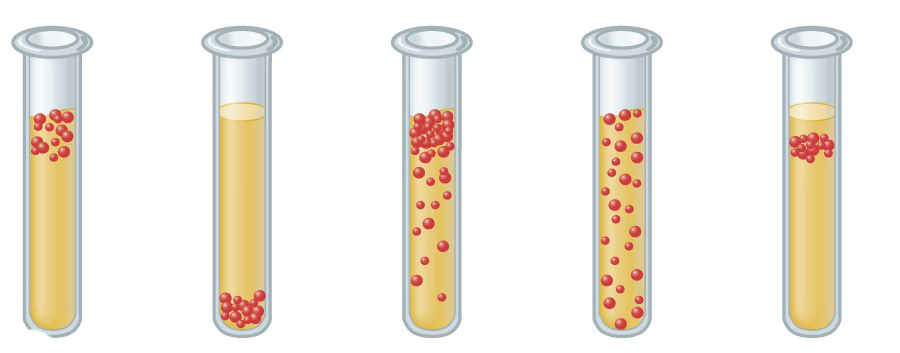
Mikroorganizmy tolerujące loterie obejmują bakterie kwasu mlekowego. Niektóre gatunki (na przykład Clostridium) mogą być odporne na wysokie stężenia tlenu ze względu na tworzenie się endospor.
Anaerobowy metabolizm energetyczny
Wszystkie beztlenowce są typowymi chemotrofami, ponieważ wykorzystują energię wiązań chemicznych jako źródło energii. W tym przypadku może być również dawcą energii materia organiczna (chemoorganotrofia) i nieorganiczna (chemolitotrofia).
Bakterie beztlenowe mają dwa rodzaje metabolizmu wolnego od tlenu: oddychanie i fermentację. Podstawowa różnica między nimi polega na mechanizmie asymilacji energii.
Tak więc, podczas fermentacji, energia jest najpierw przechowywana w postaci fosfoge- nicznej (na przykład w postaci fosfoenolopirogronianu), a następnie przy udziale dehydrogenaz cytozolowych następuje fosfanie- nie substratu ADP substratu. Elektrony są przenoszone do akceptora endogennego lub egzogennego, który staje się produktem ubocznym procesu.
Ad
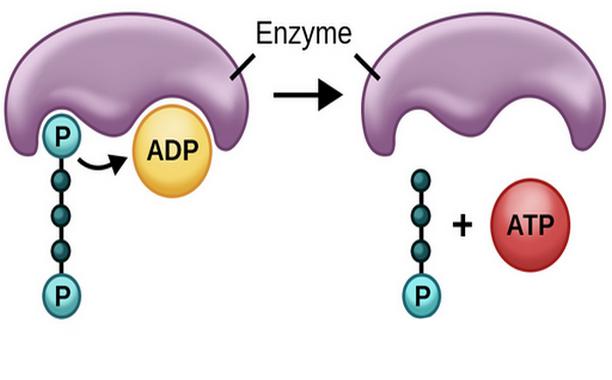
W metabolizmie typu oddechowego energia jest przechowywana w określonym związku - Pmf, który jest albo natychmiast wykorzystywany do procesów komórkowych, albo dostarczany do ukierunkowanego na błonę elektrycznego łańcucha transportowego, w którym syntetyzuje się ATP. Tylko w przeciwieństwie do oddychania tlenowego ostatecznym akceptorem elektronów nie jest tlen, ale inny związek, który może mieć zarówno charakter organiczny, jak i nieorganiczny.
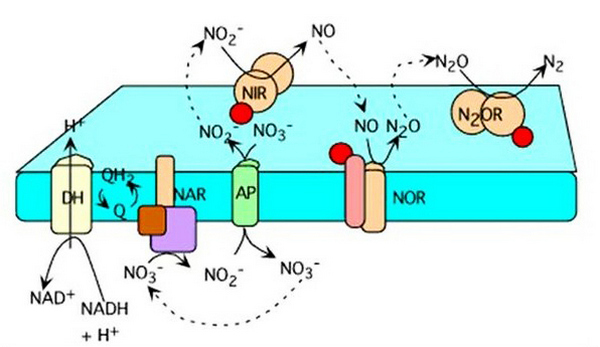
Typy beztlenowych oddechów
Głównym zadaniem, które rozwiązuje bakteria beztlenowa o typie metabolizmu oddechowego, jest znalezienie alternatywy dla tlenu cząsteczkowego. Wynik energetyczny reakcji zależy od tego. W zależności od substancji działającej jako akceptor terminalny, występują następujące rodzaje oddychania beztlenowego:
- azotan;
- żelazo;
- fumaran;
- siarczan;
- siarkowodór;
- węglan.
Oddychanie beztlenowe jest mniej skuteczne niż oddychanie tlenowe, ale w porównaniu z fermentacją daje znacznie większy wydatek energii.
Społeczność beztlenowych bakterii niszczących
Ten rodzaj mikroflory powstaje w ekologicznych niszach organicznych, w których tlen jest prawie całkowicie zużywany (zalane gleby, podziemne systemy hydrauliczne, osady mułowe itp.). Oto stopniowa degradacja związków organicznych, prowadzona przez dwie grupy bakterii:
Ad
- pierwotne bakterie beztlenowe są odpowiedzialne za pierwszy etap organicznej de-stymulacji;
- drugorzędowe beztlenowce to mikroorganizmy z metabolizmem oddechowym.
Wśród pierwotnych beztlenowców wyróżnia się hydrolitę i dyslipotrofy, które są połączone ze sobą przez oddziaływania troficzne. Hydrolityty tworzą biofilmy na powierzchni stałych substratów i produkują egzoenzymy hydrolityczne, które rozkładają złożone związki organiczne w oligomery i monomery.
Powstały substrat odżywczy jest wykorzystywany przede wszystkim przez same hydrolity, ale także przez dyslipotrofy. Te ostatnie są zwykle mniej współpracujące i nie emitują znaczących ilości egzoenzymów, pochłaniających produkty końcowej hydrolizy biopolimerów. Typowym przedstawicielem dyslipotrofów są bakterie z rodzaju Syntrophomonas.
Ad
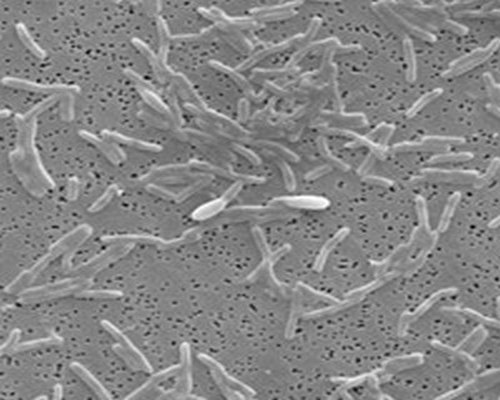
Uprawa
Specjalne wymagania dotyczące uprawy nakłada się tylko na obowiązkowe bakterie beztlenowe. Opcjonalnie dobrze reprodukuje się w środowisku tlenowym.
Metody hodowli mikroorganizmów beztlenowych można podzielić na trzy kategorie: chemiczną, fizyczną i biologiczną. Ich głównym zadaniem jest zmniejszenie lub całkowite wyeliminowanie obecności tlenu w pożywce. Stopień dopuszczalnego stężenia O 2 określa się na podstawie poziomu tolerancji określonego beztlenowego.
Metody fizyczne
Istotą fizycznych metod jest usuwanie tlenu ze środowiska powietrza, które jest w kontakcie z kulturą, lub całkowite wyeliminowanie kontaktu bakterii z powietrzem. Ta grupa obejmuje następujące technologie uprawy:
Ad
- rośnie w mikro aerostacie - specjalne urządzenie, w którym zamiast powietrza atmosferycznego powstaje sztuczna mieszanina gazów;
- głęboka uprawa - sadzenie bakterii nie na powierzchni, ale w wysokiej warstwie lub na grubość podłoża, aby powietrze tam nie przeniknęło;
- stosowanie lepkich mediów, w których dyfuzja O2 zmniejsza się wraz ze wzrostem gęstości;
- rosnące w beztlenowym słoju;
- wypełnienie powierzchni olejem wazelinowym lub parafiną;
- wykorzystanie inkubatora CO 2 ;
- wykorzystanie beztlenowej stacji SIMPLICITY 888 (najnowocześniejsza metoda).

Obowiązkową częścią metod fizycznych jest wstępne gotowanie pożywki w celu usunięcia z niej tlenu cząsteczkowego.
Używanie chemikaliów
Związki chemiczne stosowane do hodowli beztlenowców dzielą się na 2 grupy:
- Zmiatacze tlenu adsorbują cząsteczki O2 . Zdolność absorpcji zależy od rodzaju substancji i objętości przestrzeni powietrznej w środowisku. Najczęściej stosuje się pirogalol (roztwór alkaliczny), metaliczne żelazo, jednowartościowy chlorek miedzi, ditionian sodu.
- Środki redukujące (cysteina, ditiotreitol, kwas askorbinowy itp.) Zmniejszają potencjał utleniania i redukcji pożywki.
Szczególnym rodzajem metod chemicznych jest zastosowanie systemów generujących gazy, które obejmują czynniki generujące wodór i dwutlenek węgla, a O2 absorbuje katalizator palladowy. Takie systemy są używane w zamkniętych pojemnikach do uprawy (anaerostaty, plastikowe torby itp.).
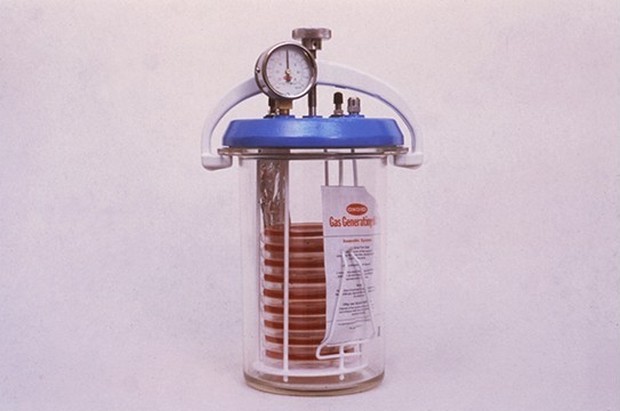
Metody biologiczne
Metody biologiczne obejmują wspólną hodowlę beztlenowców i tlenowych. Te ostatnie usuwają tlen z otoczenia, tworząc warunki dla rozwoju ich "partnerów". Jako środki pochłaniające mogą być również stosowane opcjonalne bakterie beztlenowe.
Istnieją dwie modyfikacje tej metody:
- Siew dwóch kultur na różnych połówkach szalki Petriego, która jest następnie zamykana pokrywką.
- Wysiew przy użyciu "szkiełka zegarkowego" zawierającego pożywkę z aerobową bakterią. Szkło pokrywa więc szalkę Petriego, ciągłą warstwę zaszczepioną kulturą beztlenową.
Czasami mikroorganizmy tlenowe stosuje się na etapie przygotowywania ciekłego pożywki do inokulacji beztlenowców. Po usunięciu resztkowego tlenu, tlenowce (na przykład E. colli) są zabijane przez ogrzewanie, a następnie żądana kultura jest zaszczepiana.
Izolacja czystej kultury
Czysta kultura to populacja mikroorganizmów należących do tego samego gatunku, mających takie same właściwości i uzyskanych z pojedynczej komórki. Aby uzyskać grupę bakterii o takich właściwościach, zwykle stosuje się metody rozcieńczania udaru i rozcieńczania ograniczającego, ale praca z beztlenowcami jest specjalnym procesem, który wymaga wykluczenia kontaktu z tlenem podczas otrzymywania izolowanych kolonii.
Ad
Istnieje kilka sposobów wyizolowania czystej kultury beztlenowej. Należą do nich:
- Metoda Zeisslera polega na przesiewaniu cienkiego skoku na płytkach Petriego, tworząc warunki beztlenowe, a następnie inkubację w termostacie (od 24 do 72 godzin).
- Metoda Weinberga - przydział beztlenowców w kulturze za pomocą agaru cukrowego (wysoka kolumna wysiewu), bakterie są przenoszone przez szczelną kapilarę. Najpierw materiał umieszcza się w probówce z roztworem izotonicznym (etap rozcieńczania), następnie w probówce z agarem o temperaturze 40-45 stopni, w której jest dokładnie wymieszany z podłożem. Następnie kolejne transfery przeprowadza się do 2 kolejnych probówek, z których ostatnia jest chłodzona strumieniem wody.
- Metoda Peretza - materiał rozcieńczony w izotonicznym roztworze wlewa się do szalki Petriego, tak aby wypełniała przestrzeń pod szklaną płytką na dnie, na której powinien rozpocząć się wzrost.
We wszystkich trzech metodach materiał z otrzymanych wyizolowanych kolonii przenosi się do ośrodka kontrolnego sterylności (SCS) lub podłoża Kitt-Tarozzi.