Gene Drift - czynnik ewolucji: powód, wartość, przykłady
Wraz z doborem naturalnym istnieje inny czynnik, który może wpływać na wzrost zawartości zmutowanego genu. W niektórych przypadkach może nawet wyprzeć normalną przemianę allelu. Zjawisko to nosi nazwę "dryfu genów w populacji". Rozważmy bardziej szczegółowo, czym jest ten proces i jakie są jego konsekwencje. 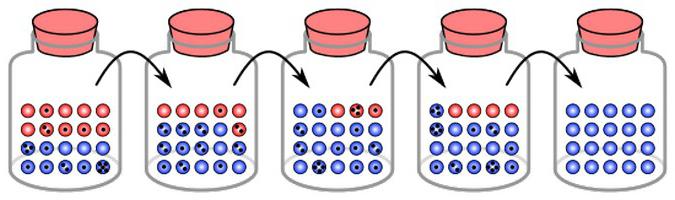
Informacje ogólne
Dryf genetyczny, którego przykłady zostaną podane w poniższym artykule, to pewne zmiany rejestrowane z pokolenia na pokolenie. Uważa się, że to zjawisko ma swoje własne mechanizmy. Niektórzy badacze obawiają się, że w puli genów wielu, jeśli nie wszystkich, narodów, ilość pojawiających się anomalnych genów rośnie obecnie dość szybko. Określają dziedziczną patologię, stanowiące warunki wstępne dla rozwoju wielu innych chorób. Uważa się również, że patomorfoza (zmiana objawów) różnych chorób, w tym chorób o charakterze mentalnym, jest właśnie dryfem genów. Omawiane zjawisko zachodzi w szybkim tempie. W rezultacie liczba zaburzenia psychiczne przyjmuje nieznane formy, stają się nierozpoznawalne w porównaniu z ich opisem w klasycznych wydaniach. Jednocześnie znaczące zmiany odnotowuje się bezpośrednio w strukturze samej choroby psychicznej. Na przykład dryf genów usuwa niektóre schizofrenii, które wcześniej występowały. Zamiast tego istnieją patologie, które trudno określić współczesnymi klasyfikatorami. 
Teoria Wrighta
Losowy dryf genów badano przy użyciu modeli matematycznych. Wykorzystując tę zasadę, Wright opracował teorię. Uważał, że kluczowe znaczenie dryftu genu w stałych warunkach obserwuje się w małych grupach. Stają się homozygotyczni, a zmienność maleje. Wright także uważał, że w wyniku zmian w grupach mogły powstać negatywne cechy dziedziczne. W rezultacie cała populacja może umrzeć bez wnoszenia wkładu rozwój gatunku. Jednocześnie selekcja odgrywa dużą rolę w wielu grupach. Pod tym względem zmienność genetyczna w populacji znów będzie nieistotna. Stopniowo grupa będzie dobrze dostosowywać się do warunków środowiskowych. Jednak kolejne zmiany ewolucyjne będą zależeć od wystąpienia korzystnych mutacji. Te procesy są dość powolne. Pod tym względem rozwój dużych populacji nie charakteryzuje się dużą prędkością. W grupach wartości pośrednich notuje się zwiększoną zmienność. Jednocześnie tworzenie nowych korzystnych genów odbywa się przez przypadek, co z kolei przyspiesza ewolucję. 
Wnioski Wrighta
Kiedy pojedynczy allel zostanie utracony z populacji, może pojawić się z powodu pewnej mutacji. Ale jeśli gatunek jest podzielony na kilka grup, w jednym z których brakuje jednego elementu, w drugim, drugi gen może migrować z miejsca, w którym się znajduje, do miejsca, w którym nie jest. Zatem zmienność pozostanie. Biorąc to pod uwagę, Wright stwierdził, że rozwój nastąpi szybciej w przypadku tych gatunków, które są podzielone na wiele różnych populacji. Jednocześnie możliwa jest między nimi migracja. Wright zgodził się z tym dobór naturalny odgrywa bardzo ważną rolę. Jednak wraz z tym wynikiem ewolucji jest dryfowanie genów. Identyfikuje trwałe zmiany w widoku. Ponadto Wright uważał, że wiele charakterystycznych cech, które powstały w wyniku dryfu, było obojętnych, aw niektórych przypadkach nawet szkodliwych dla żywotności organizmów. 
Badacze spory
Było kilka opinii na temat teorii Wrighta. Na przykład Dobzhansky uważał, że bezcelowe jest podnoszenie kwestii, który z czynników jest bardziej znaczący - dobór naturalny lub dryf genetyczny. Wyjaśnił to poprzez ich interakcję. Zasadniczo prawdopodobne są następujące sytuacje:
Ad
- Jeśli w rozwoju jednego lub innego gatunku selekcja zajmie pozycję wiodącą, zaznaczona zostanie kierunkowa zmiana w częstotliwościach genu lub stan stabilny. To ostatnie będzie zależało od warunków środowiskowych.
- Jeśli w dłuższym okresie dryf genów jest bardziej znaczący, wówczas ukierunkowane zmiany nie będą powodowane przez środowisko naturalne. Jednocześnie niekorzystne oznaki, występujące nawet w niewielkich ilościach, mogą rozprzestrzeniać się wystarczająco szeroko w grupie.
Należy jednak zauważyć, że sam proces zmiany, jak również przyczyna dryftu genów, nie jest dziś wystarczająco badany. W związku z tym nie ma jednoznacznej i konkretnej opinii na temat tego zjawiska w nauce.
Ad
Dryf genetyczny jest czynnikiem ewolucji
Z powodu zmian odnotowuje się zmianę częstotliwości alleli. Nastąpi to, dopóki nie osiągną stanu równowagi. Oznacza to, że dryf genów jest izolacją jednego elementu i utrwaleniem innego. W różnych grupach takie zmiany występują niezależnie. Pod tym względem wyniki dryfu genetycznego w różnych populacjach są różne. W końcu niektóre zestawy elementów są ustalone w niektórych, a inne w innych. Dryf genów z jednej strony prowadzi do zmniejszenia różnorodności. Równocześnie jednak powoduje również różnice między grupami, rozbieżności na niektórych podstawach. To z kolei może stanowić podstawę specjacji. 
Współczynnik wpływu
W procesie rozwoju dryf genetyczny współdziała z innymi czynnikami. Przede wszystkim nawiązuje się związek z doborem naturalnym. Współczynnik wkładu tych czynników zależy od wielu okoliczności. Przede wszystkim zależy to od intensywności selekcji. Drugą okolicznością jest wielkość grupy. Tak więc, jeśli intensywność i siła są wysokie, losowe procesy mają znikomy wpływ na dynamikę częstotliwości genetycznych. Jednocześnie w małych grupach o niewielkich różnicach w sprawności wpływ zmian jest nieporównywalnie większy. W takich przypadkach można ustalić mniej adaptacyjny allel, podczas gdy bardziej adaptacyjny zostanie utracony.
Ad
Skutki zmiany
Jednym z głównych rezultatów dryfu genetycznego jest zubożenie różnorodności w obrębie grupy. Jest to spowodowane utratą niektórych alleli i utratą innych. Z kolei proces mutacji przyczynia się do wzbogacenia różnorodności genetycznej w populacjach. Z powodu mutacji utracony allel może się powtarzać. Z uwagi na to, że dryf genetyczny jest procesem kierunkowym, jednocześnie ze zmniejszeniem zróżnicowania wewnątrzopulacji, wzrasta różnica między grupami lokalnymi. Przeciwdziałanie temu zjawisku to migracja. Tak więc, jeśli w jednej populacji allel "A" jest ustalony, w drugim - "a", to w obrębie tych grup różnorodność pojawia się ponownie. 
Wynik końcowy
Wynikiem dryfu genetycznego będzie całkowite wyeliminowanie jednego allelu i konsolidacja drugiego. Im częściej element występuje w grupie, tym większe prawdopodobieństwo jego fiksacji. Jak pokazują niektóre obliczenia, możliwość wiązania jest równa częstotliwości allelu w populacji.
Ad
Mutacje
Występują one średnio z częstotliwością 10-5 na gen na gamę na jedno pokolenie. Wszystkie allele znalezione w grupach powstały kiedyś z powodu mutacji. Im mniejsza populacja, tym mniejsze prawdopodobieństwo, że każde pokolenie będzie miało co najmniej jedną osobę - nosiciela nowej mutacji. Przy liczbie stu tysięcy każda grupa potomków z prawdopodobieństwem bliskim jedności ma zmutowany allel. Jednak jego częstotliwość w populacji, a także możliwość jego utrwalenia będzie dość niska. Prawdopodobieństwo, że ta sama mutacja pojawi się w tym samym pokoleniu, przynajmniej u jednej osoby w populacji wynoszącej 10, jest nieistotne. Jeśli jednak wystąpi w tej populacji, częstotliwość zmutowanego allelu (1 na 20 alleli), a także szanse jego utrwalenia, będą stosunkowo wysokie. W dużych populacjach pojawienie się nowego elementu występuje stosunkowo szybko. Jednocześnie jego fiksacja jest powolna. Z drugiej strony małe populacje oczekują długiej mutacji. Ale po jego wystąpieniu zapięcie szybko mija. Na tej podstawie możemy wyciągnąć następujący wniosek: szansa ustalenia neutralnych alleli zależy tylko od częstotliwości występowania mutacji. Jednocześnie wielkość populacji nie wpływa na ten proces.
Ad
Zegar molekularny
Ze względu na to, że częstotliwość występowania mutacji obojętnych u różnych gatunków jest w przybliżeniu taka sama, tempo utrwalania powinno być również w przybliżeniu równe. Wynika z tego, że liczba zmian nagromadzonych w jednym genie powinna być skorelowana z czasem niezależnej ewolucji tych gatunków. Innymi słowy, im dłuższy okres od momentu oddzielenia dwóch gatunków od jednego przodka, tym bardziej odróżniają mutacyjne substytucje. Ta zasada leży u podstaw metody zegara ewolucji molekularnej. Określa czas, jaki upłynął od momentu, w którym poprzednie generacje różnych grup systematycznych zaczęły się rozwijać niezależnie, niezależnie od siebie.
Ankieta i badanie Cukurkend
Ci dwaj amerykańscy naukowcy odkryli, że liczba różnic w sekwencji aminokwasów w cytochromie i hemoglobinie u niektórych gatunków ssaków jest wyższa, im wcześniej ich drogi ewolucyjne się rozdzieliły. Następnie wzór ten został potwierdzony dużą ilością danych eksperymentalnych. Materiał zawierał dziesiątki różnych genów i kilkaset gatunków zwierząt, mikroorganizmów i roślin. Okazało się, że przebieg godzin molekularnych odbywa się ze stałą prędkością. To odkrycie potwierdza faktyczna teoria. Zegar jest kalibrowany osobno dla każdego genu. Wynika to z faktu, że częstotliwość pojawiania się w nich neutralnych mutacji jest różna. W tym celu dokonuje się oceny liczby podstawień skumulowanych w danym genie w taksonach. Ich czas rozbieżności jest wiarygodnie ustalany na podstawie danych paleontologicznych. Po skalibrowaniu zegara molekularnego można je wykorzystać dalej. W szczególności za ich pomocą można łatwo zmierzyć czas, w którym wystąpiła rozbieżność (rozbieżność) między różnymi taksonami. Jest to możliwe, nawet jeśli ich wspólny przodek nie został jeszcze zidentyfikowany w zapisie kopalnym.