Kompleks Golgiego: opis
Kompleks Golgi został odkryty w 1898 roku. Ta struktura błony jest zaprojektowana do wydalania związków, które są syntetyzowane w retikulum endoplazmatycznym. Dalej zapoznamy się z tym systemem. 
Kompleks Golgi: budynek
Urządzenie to stos membranowych zbiorników w kształcie dysku. Te woreczki są nieco rozciągnięte w kierunku krawędzi. System bąbelkowy Golgi jest powiązany ze zbiornikami. W komórkach zwierzęcych znajduje się jeden duży lub kilka stosów, które są połączone rurami w komórki roślinne Dictyosomy są wykrywane (kilka oddzielnych stosów). Kompleks Golgi obejmuje trzy działy. Są otoczone przez bańki membranowe:
- cis - najbliżej jądra;
- środkowy;
- departament trance (najdalej od rdzenia).
Te systemy są rozróżniane przez zestaw enzymów. W dziale cis pierwsza sakwa nazywana jest "zbiornikiem ratowniczym". Przy jego pomocy receptory pochodzące z endoplazmatycznej sieci pośredniej cofają się. Oddział enzymatyczny cis o nazwie fosfoglikozydaza. Dodaje fosforan do mannozy (węglowodanów). W części środkowej znajdują się dwa enzymy. Jest to w szczególności menadiaza i transferaza N-acetyloglukozaminowa. Ten ostatni dodaje glikozoaminy. Enzymy transektora: peptydaza (prowadzi proteolizę) i transferazę (przy jej pomocy następuje transfer grup chemicznych). 
Kompleks Golgiego: funkcje
Ta struktura zapewnia rozdział białek na następujące trzy strumienie:
- Lysosomal. Według tego, białka glikowane przenikają do cis-działu aparatu Golgiego. Niektóre z nich są fosforylowane. W wyniku tego powstaje 6-fosforan mannozy - enzymy marketingososomalne. W przyszłości te fosforanowane białka wejdą do lizosomów i nie będą modyfikowane.
- Konstytutywna egzocytoza (wydzielanie). Strumień ten obejmuje białka i lipidy, które stały się składnikami urządzenia powierzchni komórki, w tym glikokaliksu. Mogą być również obecne związki, które są częścią macierzy pozakomórkowej.
- Indukowane wydzielanie. Białka, które działają poza komórką, aparat powierzchniowy, przenikają do tego strumienia w środowisku wewnętrznym w ciele. Indukowane wydzielanie jest charakterystyczne dla komórek wydzielniczych.
Zespół Golgiego bierze udział w tworzeniu wydzieliny śluzowej - mukopolisacharydów (glikozoaminoglikanów). Urządzenie to również tworzy węglowodanowe składniki glikokaliksu. Są to głównie glikolipidy. System zapewnia także siarczanowanie pierwiastków białkowych i węglowodanowych. Kompleks Golgiego bierze udział w częściowej proteolizie białek. W niektórych przypadkach związek nieaktywnego leku przechodzi do postaci aktywnej (np. Proinsulinę przekształca się w insulinę). 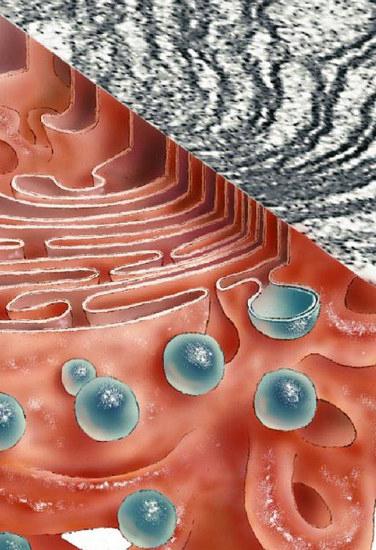
Przenoszenie związków z retikulum endoplazmatycznego (EPS)
Kompleks jest asymetryczny. Położone bliżej jądra komórki obejmują najbardziej niedojrzałe białka. Pęcherzyki, pęcherzyki błonowe są w sposób ciągły dołączane do tych torebek. Zrywają z granularnej retikulum endoplazmatycznego. Na membranach przechodzi proces syntezy białka przez rybosomy. Transport związków z retikulum endoplazmatycznego do kompleksu Golgiego jest bezkrytyczny. W tym samym czasie niepoprawnie lub niecałkowicie złożone białka pozostają w EPS. Odwrotny ruch związków do retikulum endoplazmatycznego wymaga specjalnej sekwencji sygnałowej i jest możliwy dzięki wiązaniu tych substancji z receptorami błonowymi w przedziale cis.
Modyfikacja białka
W zbiornikach kompleksu dojrzewają związki przeznaczone do wydzielania, transbłonowe, lizosomalne i inne substancje. Te białka przemieszczają się sukcesywnie wzdłuż zbiorników do organelli. Ich modyfikacje zaczynają się w nich - fosfolacja i glikozylacja. Podczas pierwszego procesu do białek przyłączona jest pozostałość. kwas fosforowy. W O-glikozylacji złożone cukry są przyłączane przez atom tlenu. Różne zbiorniki zawierają różne enzymy katalityczne. W konsekwencji następują kolejno różne procesy z dojrzewającymi w nich białkami. Niewątpliwie takie stopniowe zjawisko powinno być kontrolowane. Jako rodzaj "znaku jakości" stosowane są reszty polisacharydowe (głównie mannoza). Oznaczają białka dojrzewające. Dalsze przemieszczanie się przez zbiorniki związków nie jest w pełni zrozumiane przez naukę, pomimo faktu, że substancje oporne pozostają mniej lub bardziej związane z jedną torebką. 
Transport białek z aparatu
Z baniek pączków kompleksu. Zawierają w pełni dojrzałe związki białkowe. Główną funkcją kompleksu jest sortowanie białek przechodzących przez niego. W aparacie powstawanie "trójkierunkowego strumienia białka" - dojrzewanie i transport:
- Związki błona plazmatyczna.
- Tajemnice.
- Enzymy lizosomalne.
Poprzez transport pęcherzykowy białka, które przeszły przez kompleks Golgi, są dostarczane do określonych miejsc zgodnie z "znacznikami". Proces ten nie jest całkowicie rozumiany przez naukę. Ustalono, że transport białek z kompleksu wymaga udziału specyficznych receptorów błonowych. Rozpoznają związek i zapewniają selektywne dokowanie pęcherzyka i jednego lub drugiego organellum. 
Tworzenie się lizosomu
Przez aparat przechodzi wiele enzymów hydrolitycznych. Dodatek wspomnianej powyżej znacznika jest przeprowadzany przy udziale dwóch enzymów. Specyficzne rozpoznawanie hydrolaz lizosomalnych przez pierwiastki ich trzeciorzędowej struktury i dodanie fosforanu N-acetyloglukozaminy przeprowadza się za pomocą fosfotransferazy N-acetyloglukozaminy. Fosfoglikozyd, drugi enzym, usuwa N-acetyloglukozaminę, w wyniku czego powstaje etykieta M6F. To z kolei jest rozpoznawane przez białko receptorowe. Przy jego pomocy hydrolazy są dostarczane do pęcherzyków i ich mandatów do lizosomów. W kwaśnych warunkach fosforan jest odszczepiany od dojrzałej hydrolazy. Jeżeli występują zakłócenia w aktywności fosfotransferazy N-acetyloglukozaminy z powodu mutacji lub z powodu defektów genetycznych w receptorze M6F, wszystkie enzymy lizosomalne są dostarczane domyślnie do błony zewnętrznej. Następnie są wydzielane do warunków pozakomórkowych. Ustalono także, że niektóre z receptorów M6P są również transportowane do błony zewnętrznej. Dokonują zwrotu losowo złapanych enzymów lizosomalnych ze środowiska zewnętrznego wewnątrz komórki podczas endocytozy. 
Transport substancji do błony zewnętrznej
Zwykle nawet na etapie syntezy białkowe związki błony zewnętrznej z ich hydrofobowymi regionami są osadzone w ścianie retikulum endoplazmatycznego. Następnie są dostarczane do kompleksu Golgi. Stamtąd są transportowane na powierzchnię komórki. W procesie fuzji plazmalemy i pęcherzyków takie związki nie są uwalniane do środowiska.
Sekrecja
Prawie wszystkie związki wytwarzane w komórce (o charakterze zarówno białkowym, jak i niebiałkowym) przechodzą przez kompleks Golgiego. Tam rozwijają się w pęcherzyki wydzielnicze. W roślinach z dictyos materiał jest w ten sposób wytwarzany. ściana komórkowa.