Czym jest chromatyna? Funkcje chromatyny
Materiał genetyczny organizmów eukariotycznych ma bardzo złożoną organizację. Cząsteczki DNA, znajdujące się w jądrze komórkowym, są częścią specjalnej wieloskładnikowej substancji - chromatyny.
Definicja pojęcia
Chromatyna jest materiałem jądra komórkowego zawierającego informację dziedziczną, która jest złożonym funkcjonalnym kompleksem DNA z białkami strukturalnymi i innymi elementami, które zapewniają pakowanie, przechowywanie i realizację genomu karyiotic. W uproszczonej interpretacji jest to substancja, która tworzy chromosomy. Termin pochodzi od greckiego "chrome" - kolor, farba.
Koncepcja została wprowadzona przez Fleminga w 1880 roku, ale wciąż trwa debata na temat tego, czym jest chromatyna z punktu widzenia jej składu biochemicznego. Niepewność dotyczy niewielkiej części składników, które nie biorą udziału w tworzeniu struktury cząsteczek genetycznych (niektóre enzymy i kwasy rybonukleinowe).
Ad
Na elektronicznym zdjęciu jądra międzyfazowego chromatynę wizualizuje się jako liczne płaty ciemnej materii, które mogą być małe i rozproszone lub połączone w duże gęste skupiska.
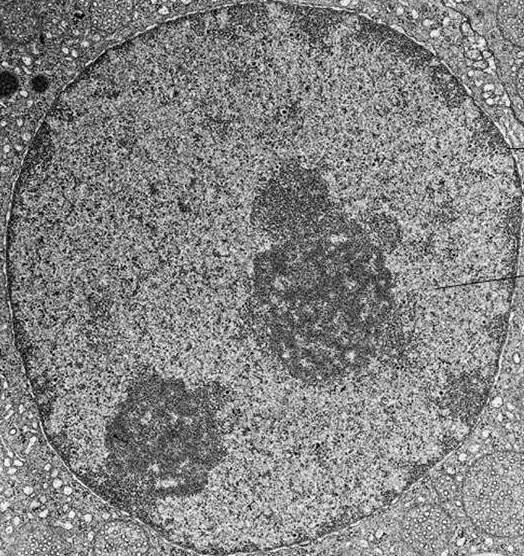
Kondensacja chromatyny podczas podziału komórki prowadzi do powstania chromosomów, które są widoczne nawet w konwencjonalnym mikroskopie świetlnym.
Strukturalne i funkcjonalne składniki chromatyny
Aby określić, jaka chromatyna jest na poziomie biochemicznym, naukowcy wyodrębnili tę substancję z komórek, przenieśli ją do roztworu i zbadali skład i strukturę tego składnika w tej postaci. Zastosowano zarówno metody chemiczne, jak i fizyczne, w tym technologie mikroskopii elektronowej. Okazało się, że skład chemiczny chromatyny o 40% jest reprezentowany przez długie cząsteczki DNA i prawie 60% przez różne białka. Te ostatnie są podzielone na dwie grupy: histonów i nie-histonów.
Ad
Histony to duża rodzina rdzeniowych białek jądrowych, które wiążą się silnie z DNA, tworząc strukturalny szkielet chromatyny. Ich liczba jest w przybliżeniu równa procentowi cząsteczek genetycznych.
Reszta (do 20%) frakcji białkowej przypada na białka wiążące DNA i modyfikujące przestrzenność, a także na enzymy biorące udział w procesie odczytu i kopiowania informacji genetycznej.
Oprócz głównych elementów, w składzie chromatyny kwasów rybonukleinowych (RNA), glikoproteiny, węglowodany i lipidy znajdują się w niewielkiej ilości, ale kwestia ich powiązania z kompleksem pakowania DNA jest wciąż otwarta.
Histony i nukleosomy
Masa cząsteczkowa histonów waha się od 11 do 21 kDa. Duża liczba zasadowych aminokwasów, lizyny i argininy, nadaje tym białkom ładunek dodatni, przyczyniając się do tworzenia wiązań jonowych z przeciwnie naładowanymi grupami fosforanowymi podwójnej helisy DNA.
Istnieje 5 typów histonów: H2A, H2B, H3, H4 i H1. Pierwsze cztery typy biorą udział w tworzeniu głównej jednostki strukturalnej chromatyny - nukleosomu, który składa się z rdzenia (rdzenia białka) i owiniętego wokół niego DNA.
Rdzeń nukleosomalny jest reprezentowany przez ośmio-cząsteczkowy oktamerowy kompleks histonów, który obejmuje tetramer H3-H4 i dimer H2A-H2B. Region DNA o długości około 146 par nukleotydowych jest nawijany na powierzchnię cząsteczki białka, tworząc 1,75 cewki i przechodzi do sekwencji łącznikowej (około 60 bp) łączącej nukleosomy ze sobą. Cząsteczka H1 wiąże się z DNA linkera, chroniąc go przed działaniem nukleaz.
Ad
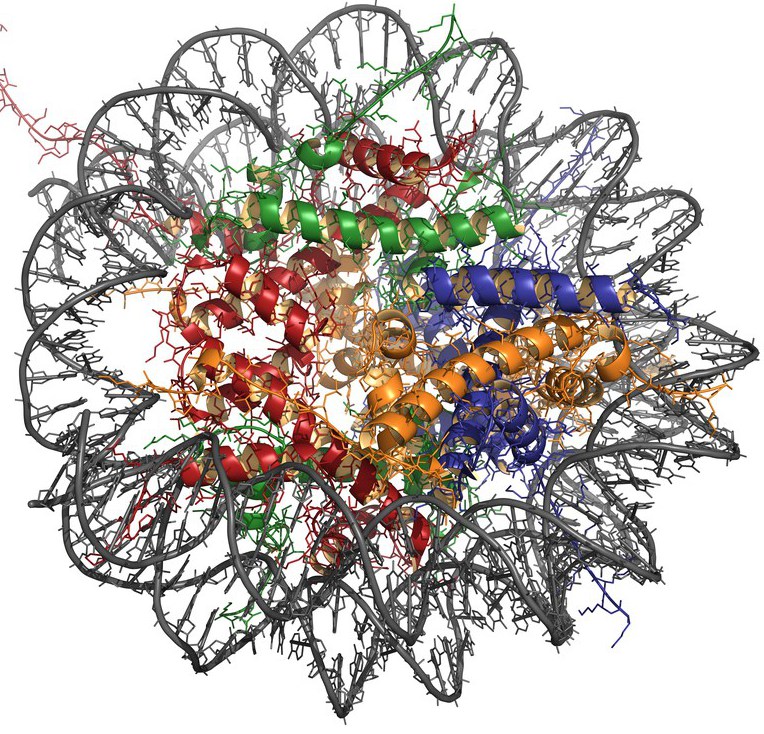
Histony mogą ulegać różnym modyfikacjom, takim jak acetylacja, metylacja, fosforylacja, ADP-rybozylacja i interakcja z białkiem ubikwityny. Te procesy wpływają na przestrzenną konfigurację i gęstość pakowania DNA.
Niehistonowe białka
Istnieje kilkaset odmian niehistonowych białek o różnych właściwościach i funkcjach. Ich masa cząsteczkowa waha się od 5 do 200 kDa. Specjalna grupa składa się z białek specyficznych dla miejsca, z których każda jest komplementarna do określonego regionu DNA. Ta grupa obejmuje 2 rodziny:
- "palce cynkowe" - rozpoznają fragmenty 5 par nukleotydów;
- homodimery charakteryzują się strukturą spiro-skręt-helisa w fragmencie związanym z DNA.
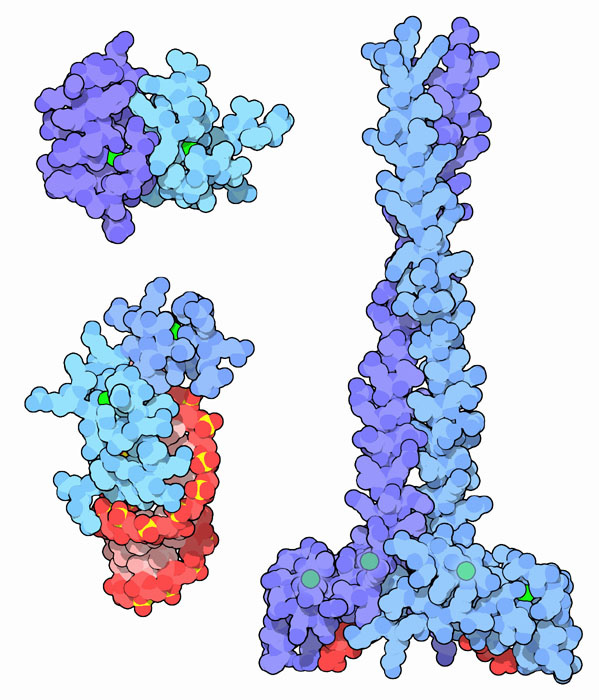
Najlepiej badane są tak zwane białka o wysokiej ruchliwości (białka HGM), które są trwale związane z chromatyną. Ta nazwa rodziny została odebrana ze względu na dużą prędkość ruchu cząsteczek białek w żelu elektroforetycznym. Ta grupa zajmuje większość frakcji nie-histonowej i obejmuje cztery główne typy białek HGM: HGM-1, HGM-14, HGM-17 i HMO-2. Wykonują funkcje strukturalne i regulacyjne.
Niehormonowe białka obejmują również enzymy, które zapewniają transkrypcję (syntezę informacyjnego RNA), replikację (podwojenie DNA) i naprawę (naprawa uszkodzeń w cząsteczce genetycznej).
Poziom zagęszczenia DNA
Specyficzność struktury chromatyny jest taka, że umożliwia ona nici DNA o długości całkowitej większej niż metr, aby pasowały do jądra o średnicy około 10 mikronów. Jest to możliwe dzięki wielostopniowemu systemowi opakowań cząsteczek genetycznych. Ogólny schemat zagęszczania obejmuje pięć poziomów:
Ad
- włókno nukleosomowe o średnicy 10-11 nm;
- fibryl 25-30 nm;
- domeny pętlowe (300 nm);
- Włókno o grubości 700 nm;
- chromosomy (1200 nm).
Ta forma organizacji zapewnia redukcję długości pierwotnej cząsteczki DNA o 10 tysięcy razy.
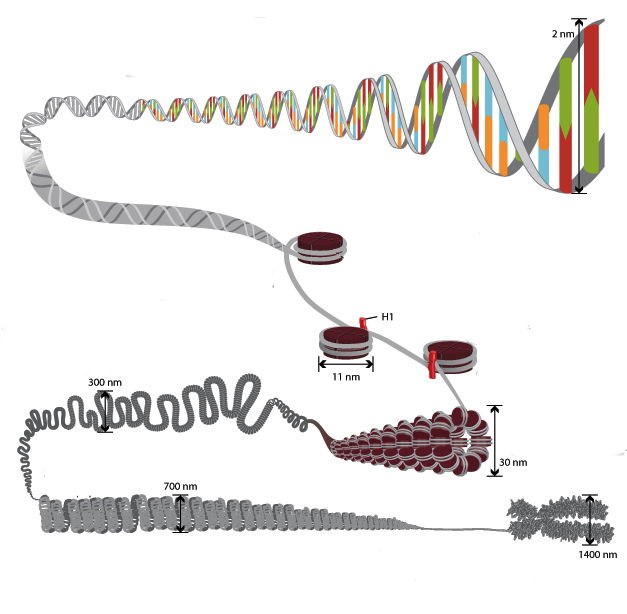
Wątek o długości 11 nm jest utworzony przez szereg nukleosomów połączonych regionami DNA linkera. Na mikroskopie elektronowym taka struktura przypomina korale nawleczone na żyłce. Jądro nukleosomalne składa się w cewkę podobną do solenoidu, tworząc włóknistą warstwę o grubości 30 nm. Histon H1 bierze udział w jego tworzeniu.
Włókno solenoidowe składa się w pętle (czyli domeny), które są przymocowane do wspierającej macierzy międzyjądrowej. Każda domena zawiera od 30 do 100 tysięcy par nukleotydów. Ten poziom zagęszczenia jest charakterystyczny dla chromatyny międzyfazowej.
Struktura o grubości 700 nm jest uformowana przez spiralowanie włókienka domeny i jest nazywana chromatydem. Z kolei dwie chromatydy tworzą piąty poziom organizacji DNA - chromosom o średnicy 1400 nm, który staje się widoczny na etapie mitozy lub mejozy.
Ad
Tak więc chromatyna i chromosom są formami opakowań dla materiału genetycznego, które zależą od cyklu życiowego komórki.
Chromosomy
Chromosom składa się z dwóch siostrzanych chromatyd identycznych ze sobą, z których każda jest utworzona przez jedną superskręconą cząsteczkę DNA. Połówki połączone są specjalnym włóknistym ciałem, zwanym centromerem. W tym samym czasie struktura ta jest zwężeniem, które oddziela każdą chromatydę od ramion.
W przeciwieństwie do chromatyny, która jest materiałem strukturalnym, chromosom jest odrębną jednostką funkcjonalną, charakteryzującą się nie tylko strukturą i kompozycją, ale także unikalnym zestawem genetycznym, a także pewną rolą we wdrażaniu mechanizmów dziedziczności i zmienności na poziomie komórkowym.
Euchromatyna i heterochromatyna
Chromatyna w jądrze występuje w dwóch postaciach: mniej spiralizująca (euchromatyna) i bardziej zwarta (heterochromatyna). Pierwsza forma odpowiada transkrypcyjnie aktywnym odcinkom DNA, a zatem nie jest tak mocno ustrukturyzowana. Heterochromatyna jest podzielona na opcjonalne (może przechodzić od aktywnej do gęstej nieaktywnej formy w zależności od etapu cyklu życia komórki i potrzeby wdrożenia pewnych genów) i konstytutywnej (stale skondensowanej). Podczas podziału mitotycznego lub mejotycznego cała chromatyna jest nieaktywna.
Konstytutywną heterochromatynę znaleziono w pobliżu centromerów i w końcowych regionach chromosomu. Wyniki mikroskopii elektronowej pokazują, że taka chromatyna zachowuje wysoki stopień kondensacji nie tylko na etapie podziału komórki, ale także podczas fazy międzyfazowej.
Biologiczna rola chromatyny
Główna funkcja chromatyny jest gęsto upakowana dużą ilością materiału genetycznego. Jednak samo umieszczenie DNA w jądrze nie jest wystarczające do funkcjonowania komórki. Konieczne jest, aby te cząsteczki właściwie "działały", to znaczy, mogły przenosić zawarte w nich informacje za pośrednictwem układu DNA-RNA-białko. Ponadto komórka musi rozprowadzać materiał genetyczny podczas podziału.
Urządzenie chromatyny w pełni spełnia te cele. Część białkowa zawiera wszystkie niezbędne enzymy, a specyfika struktury pozwala im wchodzić w interakcje z pewnymi fragmentami DNA. Dlatego drugą ważną funkcją chromatyny jest zapewnienie wszystkich procesów związanych z implementacją genomu jądrowego.