Podwójne nawożenie w roślinach
Podwójne nawożenie roślin ma wielkie znaczenie biologiczne. Zostało odkryte przez Navashin w 1898 roku. Następnie przyjrzymy się bliżej, jak podwójne nawożenie występuje w roślinach. 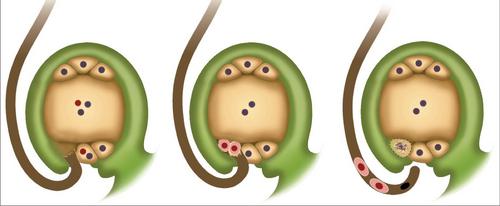
Znaczenie biologiczne
Proces podwójnego nawożenia sprzyja aktywnemu rozwojowi tkanki odżywczej. W związku z tym, ovule nie przechowuje materiału na przyszłość. To z kolei wyjaśnia jego szybki rozwój.
Podwójny schemat nawożenia
Krótko mówiąc, zjawisko to można opisać następująco. Podwójne zapłodnienie w okrytozalążku polega na wniknięciu dwóch plemników do jajnika. Jeden łączy się z jajkiem. Przyczynia się to do rozpoczęcia rozwoju diploidalnego zarodka. Drugi nasienie łączy się z centralną komórką. W wyniku tego powstaje element triploidalny. Biel pochodzi z tej komórki. Jest to odżywczy materiał dla rozwijającego się zarodka.
Ad
Budowa probówki pyłkowej
Podwójne zapłodnienie okrytonasiennych zaczyna się po wytworzeniu wysoce zredukowanego generowania haploidów. Jest reprezentowana przez gametofity. Podwójne nawożenie roślin kwiatowych sprzyja kiełkowaniu pyłków. Rozpoczyna się od pęcznienia ziarna i następującego po nim powstawania łagiewki pyłkowej. Przełamuje sporoderm w najcieńszej części. Nazywa się to aperturą. Z końcówki łagiewki emitowane są określone substancje. Zmiękczają tkanki w kolumnie i stygmatyzacji. Z tego powodu zawierają one łagiewkę pyłkową. Gdy rozwija się i rośnie, zarówno plemniki, jak i jądro z komórki wegetatywnej przechodzą do niego. W przeważającej większości przypadków, łagiewka pyłkowa przenika do jądra (megasporangia) poprzez mikroplazję zalążka. Zdarza się to rzadko w inny sposób. Po wniknięciu do torebki zarodkowej pyłek pęka. W rezultacie cała jego zawartość wlewa się do środka. Podwójne nawożenie roślin kwiatowych kontynuuje się z utworzeniem diploidalnej zygoty. To przyczynia się do pierwszego plemnika. Drugi element jest połączony z wtórnym jądrem, które znajduje się w centralnej części worka zarodkowego. Utworzone jądro triploidalne przekształca się następnie w bielmo. 
Tworzenie się komórek: informacje ogólne
Proces podwójnego nawożenia roślin kwitnących odbywa się za pomocą specjalnych komórek rozrodczych. Ich powstawanie odbywa się w dwóch etapach. Pierwszy etap nazywa się sporogenezą, drugi - hematogenezą. W przypadku tworzenia się komórek męskich etapy te określa się jako mikrosporogenezę i mikrohematogenezę. Podczas formowania się żeńskich elementów narządów płciowych przedrostek zmienia się na "mega" (lub "makro"). Sporogeneza opiera się na mejozie. Jest to proces tworzenia haploidalnych elementów. Mejoza, a także wśród przedstawicieli fauny, poprzedza rozmnożenie komórek przez podziały mitotyczne.
Ad
Produkcja nasienia
Pierwotne formowanie męskich narządów płciowych odbywa się w specjalnej tkance pylnika. Nazywa się archesparial. W nim, w wyniku działania mitoz, pojawia się powstawanie licznych elementów, komórek pyłkowych matki. Następnie wchodzą w mejozę. Z powodu dwóch podziałów mejotycznych powstają 4 haploidalne mikrospory. Przez jakiś czas leżą w pobliżu, tworząc tetrady. Następnie rozpadają się na ziarna pyłku - pojedyncze mikrospory. Każdy z uformowanych elementów zaczyna być pokryty dwiema powłokami: zewnętrzną (wewnętrzną) i wewnętrzną (wewnętrzną). Następnie rozpoczyna się kolejny etap - mikrogametogeneza. To z kolei składa się z dwóch kolejnych podziałów mitotycznych. Po utworzeniu pierwszych dwóch komórek: generatywnych i wegetatywnych. Następnie pierwszy przekazuje kolejny podział. W rezultacie powstają dwie męskie komórki - plemniki. 
Makroporogeneza i megasporogeneza
W tkankach jajowodu jeden lub więcej elementów archespery zaczyna się rozdzielać. Zaczynają szybko rosnąć. Dzięki tej aktywności stają się znacznie większe niż pozostałe komórki otaczające je w zalążku. Każdy element arpspore jest dzielony przez mitozę jeden, dwa lub więcej razy. W niektórych przypadkach komórka może natychmiast przekształcić się w matkę. Mejoza pojawia się w środku. W rezultacie powstają 4 komórki haploidalne. Z reguły największy z nich zaczyna się rozwijać, zamieniając się w torebkę zarodkową. Trzy pozostałe stopniowo degenerują się. Na tym etapie makrosporogeneza zostaje zakończona, rozpoczyna się hematogeneza makr. Podczas tego procesu występują podziały mitotyczne (większość roślin okrytonasiennych ma trzy). Cytokinezy nie towarzyszą mitozy. W wyniku trzech podziałów powstaje worek zarodkowy z ośmioma jądrami. Następnie izolują się w niezależne komórki. Te elementy są rozprowadzane w pewien sposób przez torebkę materiału biologicznego. Jedna z izolowanych komórek, która w rzeczywistości jest komórką jajową, wraz z dwoma innymi synergistami, zachodzi na mikropolu, do którego dochodzi do penetracji plemników. W tym procesie bardzo ważną rolę odgrywają synergetyki. Zawierają enzymy, które przyczyniają się do rozpuszczania błon na łagiewkach pyłkowych. Po przeciwnej stronie worka zarodkowego znajdują się pozostałe trzy komórki. Nazywa się je antypodami. Za pomocą tych pierwiastków składniki odżywcze są przenoszone z jaj do worka biologicznego. Pozostałe dwie komórki znajdują się w części środkowej. Często się łączą. W wyniku ich połączenia powstaje diploidalna centralna komórka. Po podwójnym zapłodnieniu i przeniknięciu plemników do jajnika, jedno z nich, jak wspomniano powyżej, połączy się z komórką jajową. 
Wyposażony w łagiewkę pyłkową
Podwójnemu nawożeniu towarzyszy jego interakcja z tkankami sporofitu. Jest to dość specyficzne. Proces ten jest regulowany przez aktywność związków chemicznych. Stwierdzono, że pyłek jest płukany woda destylowana ona straci zdolność kiełkowania. Jeśli otrzymany roztwór zostanie zatężony, a następnie przetworzony, wówczas ponownie stanie się kompletny. Rozwój łagiewki pyłkowej po kiełkowaniu kontroluje tkankę słupka. Na przykład w przypadku bawełny jego wzrost do komórki jajowej zajmuje około 12-18 godzin. Jednak już po 6 godzinach możliwe jest ustalenie, do której zalążek pójdzie pyłek. Jest to zrozumiałe, ponieważ zaczyna się w nim niszczenie synergii. Obecnie nie ustalono, w jaki sposób roślina może kierować rozwojem rury we właściwym kierunku oraz w jaki sposób synergetyk dowiaduje się o przybliżeniu. 
"Zakaz" na selfing
Jest często obserwowany w roślinach kwitnących. Zjawisko to ma swoją własną charakterystykę. "Zakaz" samozapylenia przejawia się w tym, że sporofit "identyfikuje" swojego własnego męskiego hematofitę i nie pozwala mu uczestniczyć w zapłodnieniu. Jednocześnie w niektórych przypadkach kiełkowanie własnego pyłku nie występuje na stygnie słupka. Jednak z reguły wzrost rurki nadal się rozpoczyna, ale później zatrzymuje się. W rezultacie pyłek nie dociera do jaja, w wyniku czego nie występuje podwójne nawożenie. Darwin zauważył to zjawisko. Tak więc znalazł w pierwiosnce wiosenne kwiaty dwie formy. Niektóre z nich miały długie łodygi z krótkimi pręcikami. Inne mają krótką kolumnę. W nich filamenty staminate były długie. Rośliny o krótkich łodygach różnią się dużym pyłkiem (dwukrotnie większym niż w innych). W tym samym czasie komórki w brodawkach stygmatu są małe. Te objawy są kontrolowane przez grupę ściśle utkanych genów. 
Receptory
Podwójne nawożenie jest skuteczne, gdy pyłek przenosi się z jednej formy do drugiej. Do rozpoznania własnych elementów należą odpowiedzialne specjalne cząsteczki receptorowe. Są to złożone związki węglowodanów z białkami. Ustalono, że formy dzikiej kapusty, które nie wytwarzają tych cząsteczek receptora w tkankach stygmatyzacji, mogą ulegać samozapyleniu. Dla normalnych roślin związki węglowodanów-białek pojawiają się dzień przed otwarciem kwiatu. Jeśli otworzysz pączek i przetworzysz go z własnym pyłkiem dwa dni przed jego kwitnieniem, nastąpi podwójne nawożenie. Jeśli zostanie to zrobione dzień przed otwarciem, nie będzie.
Ad
Allele
Warto zauważyć, że w niektórych przypadkach "samo-niekompatybilność" pyłku w roślinach jest ustalana za pomocą szeregu wielu elementów pojedynczego genu. Zjawisko to jest podobne do niekompatybilności transplantacji tkanek u zwierząt. Takie allele są oznaczone literą S. Liczba w populacji tych elementów może osiągnąć dziesiątki, a nawet setki. Na przykład, jeśli genotyp rośliny wytwarzającej jaja to s1s2, a s2s3 wytwarzający pyłek, podczas zapylenia krzyżowego, kiełkowanie zostanie odnotowane tylko w 50% cząstek pyłu. Będą to ci, którzy niosą allel s3. Jeśli istnieje kilka tuzinów elementów, wówczas większość pyłków kiełkuje normalnie podczas zapylenia krzyżowego, podczas gdy samozapylenie jest całkowicie uniemożliwione. 
Podsumowując
W przeciwieństwie do nagonasiennych, które charakteryzują się rozwojem wystarczająco silnego haploidalnego endospermy, niezależnie od zapłodnienia, w okrytozalążkowych tkankach tworzy się tylko w tym jednym przypadku. Biorąc pod uwagę ogromną liczbę generacji, osiągnięto znaczne oszczędności energii. Zwiększenie stopnia ploidalności bielma najwyraźniej przyczynia się do szybszego wzrostu tkanki w porównaniu z diploidalnymi warstwami sporofitu.